Natural historians have known for a long time that islands often harbour an extraordinary amount of biodiversity. One reason is that lineages that colonize islands can exploit open ecological niches and therefore diversify along new evolutionary trajectories. So did Anolis lizards following their arrival to the Caribbean. A range of habitat specialists has evolved repeatedly on different islands, creating a structured diversity of morphologies. Since one Anolis clade has remained on the mainland, and one clade has returned the mainland, this group of lizards presents a kind of before/after ‘island evolution’ experimental setting that can help us understand how evolution works.
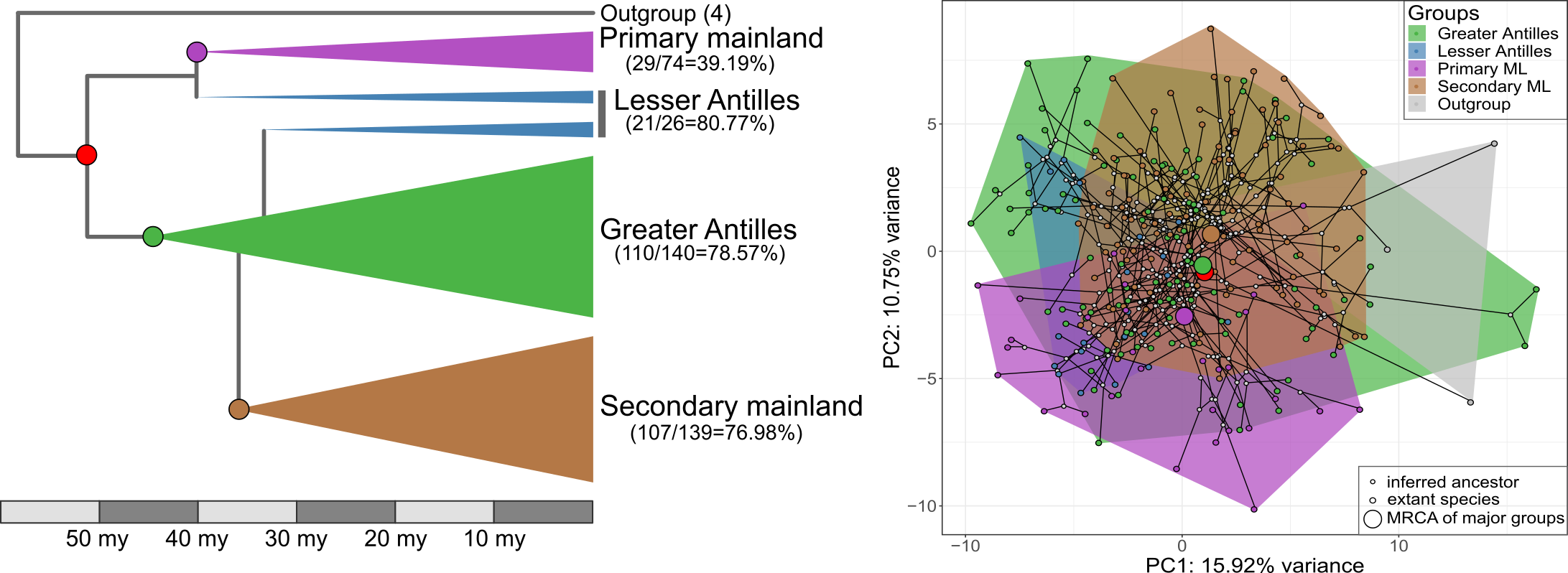
Left: Phylogenetic relationship between major groups of Anolis lizards. Right: The first and third PCs color-coded by biogeographic group.
Our new study, published in Nature Communications, took advantage of this setting to find out if morphological diversification proceeded in a different way following colonization of the Caribbean islands compared to the lizards that stayed on the mainland. This would be predicted if there were lots of ecological opportunities on islands. Furthermore, we wanted to know if the clade that returned to the mainland evolved along similar trajectories as their distant mainland relatives, or if they followed more closely their Caribbean relatives. Finally, we also wanted to test if evolution on the Caribbean islands was faster, more punctuated, and led to higher morphological diversity than on the mainland, again expected if lizards encountered a lot of empty niches on islands.
To answer these questions, Nathalie and Illiam travelled to the Nanoscale Research Facility at the University of Florida to collect CT-scans of more than 700 individuals of 271 Anolis species. Back home they quantified the morphology of the parts of the skeleton that determine how the lizards navigate their habitat – limb bones, hip and shoulder girdles.
Interestingly, we found that the evolutionary modularity – the covariation of parts of the locomotor skeleton across the evolutionary tree – was different between the clades: while the lizards whose ancestors had never set foot on an island showed a tight covariation between the fore- and hindlimbs, Anolis evolution on islands followed a trajectory with much tighter covariation between limbs and their respective girdles (forming a front and a hind module). What is more, the evolutionary modularity that was seen across island lizards was also seen in the mainland re-colonizers.
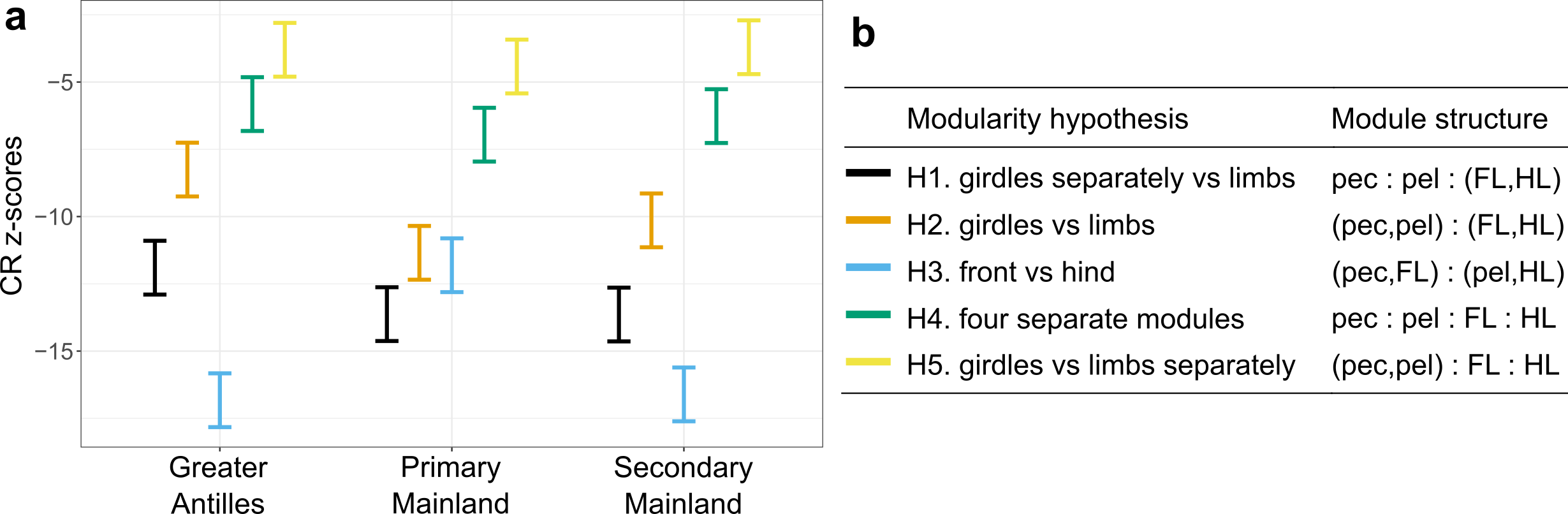
Left: CR z-scores (effect sizes) of the five alternative modularity hypotheses, evaluated separately for the three major groups. Right: Configurations of the five alternative modularity hypotheses.
What does this mean? The answers are hidden in the past, of course, but we speculate that the exploitation of ecological opportunities following colonization of the islands led to a profound change in how limbs and girdles develop and grow together during ontogeny. That is, adaptation on islands may have changed how limbs and girdles vary together, and this change persisted even after lizards recolonized the mainland. This meant that lizards in the clade that recolonized the mainland produced very similar morphologies to those seen on the islands, but failed to explore some parts of the morphospace occupied by their other mainland relatives.
If we compare the island lizards with the clade that returned to the mainland, we find that a general prediction from island biogeography holds true – island lizards show faster and more variable evolutionary rates and greater morphological variation. That is, while the patterns of diversification of the locomotor skeleton is similar, the mainland lizards produce only a subset of the morphologies seen on the islands. The same is not true, however, if we compare the morphology of island lizards to the morphology of lizards in the ‘primary’ mainland clade. This mainland clade is equally diverse as the lizards on islands, and it contains morphologies that are not seen on the islands. Perhaps this means that comparing diversification patterns of island and mainland groups is only ‘fair’ if the two groups share the same variability. Or perhaps it reflects historical differences in the ecological opportunities for adaptive diversification in the two mainland clades.
 From left to right, Anolis christophei, Anolis porcus and Anolis rejectus
From left to right, Anolis christophei, Anolis porcus and Anolis rejectus
We do not yet have all the answers, and many will forever remain unknown. However, we do suggest that these macroevolutionary trends of the locomotor skeleton in Anolis illustrate that adaptation to ecological opportunities on islands can have profound effects on trait development, with lasting effects on patterns of morphological diversification. To put this hypothesis to the test, we need to study how the parts of the locomotor skeleton vary together within species, and do this for several species across the different clades. A big task, but not impossible.
This post was originally published here at the Uller Lab blog page.
Read the publication here: Feiner, N., Jackson, I.S.C., Stanley, E.L. et al. Evolution of the locomotor skeleton in Anolis lizards reflects the interplay between ecological opportunity and phylogenetic inertia. Nat Commun 12, 1525 (2021). https://doi.org/10.1038/s41467-021-21757-5
Sofia Casasa, a recent graduate student under the EES research program, is now a postdoctoral researcher at the Ragsdale lab at Indiana University. We are delighted to cross-post Dr. Casasa’s recent “behind the paper” article (from the Nature Research Ecology & Evolution Community) on her new paper “What can horned beetles tell us about the mechanisms of plasticity and their evolution?”
We also interviewed Dr. Casasa to learn more about her work and the unique challenges that come with the empirical study of plasticity. Find out what she’ll be working on next at the end of the post!
What can horned beetles tell us about the mechanisms of plasticity and their evolution?
How do organisms interact with their environments? Research on phenotypic plasticity focuses on how organisms with the same genotype can adjust their phenotypes in response to different environmental conditions. Plasticity can take many forms and can occur at any stage during development, but those changes that occur early in development are commonly irreversible and it is those changes that fall under the rubric of developmental plasticity.
An extreme case of developmental plasticity are polyphenisms, where instead of showing a gradual response to a particular environmental factor, individuals develop into distinctly different, alternative phenotypes. One of the most well-known examples of polyphenism are castes in social insects (bees, ants, etc.), in which nutritional conditions determine whether females with the same genotype develop into workers or queens, with no intermediates.
Yet, what are the mechanisms that specify development of one or the other phenotype? And, how does plasticity itself evolve? Are the genetic mechanisms underlying plasticity responsible for the different degrees of plasticity seen across species?

Three species of horned beetles used: Digitonthophagus gazella, Onthophagus taurus and Onthophagus sagittarius (from left to right).
I have been tackling these questions for the past several years. Our recently published article in Nature Ecology and Evolution was the last chapter of my PhD dissertation, and tried to answer some of these questions from a comparative transcriptomic approach. My advisor, Armin Moczek, and I decided to collaborate with Eduardo Zattara, a former postdoc in the lab who has ample experience in analyzing transcriptomes. Despite having returned to Argentina, Eduardo was the perfect mentor to remotely guide me through all the bioinformatic tools needed for this project.
To address our questions, we focused on horned dung beetles from the tribe Onthophagini. In these beetles, the quality and quantity of nutrition received during larval development dictates whether males develop as large individuals with large horns or as small individuals with no horns. Large horns are used in male-male combat to gain access to females, while hornless males rely on more sneaky tactics to circumvent large males and intercept females. Importantly, like the castes of social insects horned and hornless males, too, constitute a polyphenism with very few intermediates seen in nature.
The wide diversity of horns and plastic responses found among Onthophagini allowed us to explore the mechanisms and evolution of plasticity. We focused on three species of horned beetles that show vast differences in the degree of plasticity: Onthophagus taurus, which exhibits an extreme, explosive response to nutrition; closely related Onthophagus sagittarius, which has secondarily lost horns; and distantly related Digitonthophagus gazella, which displays a modest plastic response, similar to what it is thought to be the ancestral condition for the tribe.

Left: Cattle farm where beetles and dung are regularly collected. Right: 12-well plate with D. gazella larvae
To collect tissues for transcriptomic analyses the first step is to get adult beetles from the field to start beetle colonies. Beetles are collected in organic cattle farms by our collaborators in North Carolina (O. taurus) and Hawaii (O. sagittarius and D. gazella), and then shipped to Bloomington, Indiana, where we keep them in separate colonies of ~300 beetles. Females build dung balls and lay a single egg in each ball. We collect the balls and open them to move the recently hatched larvae to be artificially reared in 12-well plates. This set up allowed us to manipulate nutrition levels by supplying dung of varying quality and pick either large, horned or small, hornless individuals of each species that were later used to dissect horn tissue and perform RNA extractions and sequencing.
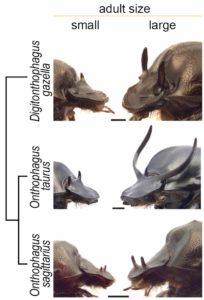
Small, low-nutrition males (left) and large, high-nutrition males (right) of the three species of beetle used in this study and their phylogenetic relationship.
We were particularly interested in understanding the relationship between gene expression changes across nutrition conditions and what we saw at the morphological level (horned vs hornless). Are differences in gene expression between small and large individuals greater in the species showing the more dramatic morphological response? How about the species that has lost the plastic response? Using RNAseq to estimate genome-wide expression in horn tissues, we found that gene expression differences mirrored what we saw at the morphological level; more exaggerated polyphenisms correspond with an increased number of nutrition-responsive genes. While this may seem intuitive, there was no previous evidence or expectation that this would be the case (e.g. we could have seen the same number of genes but more growth promoters vs. inhibitors in extreme responses).
Finally, we explored several additional questions that remain heavily debated in the field. For example, genetic accommodation is a process proposed to explain how variation in plastic responses among individuals can fuel evolutionary diversification of plasticity among species. Our data is consistent with such a scenario: ancestral plasticity in gene expression may have contributed to the evolution of growth exaggeration as well as loss in the Onthophagini tribe. Similarly, whether the evolution of novel degrees of plasticity is made possible by new genes (so called taxon-restricted genes) or facilitated by already existing, conserved, and re-purposed genes was unknown. We found evidence that a combination of both contributed to the exaggeration and loss of plasticity. As such our work advances our understanding of the evolution of developmental plasticity, a fundamental feature of most forms of life on our planet.
This blog post has been reproduced with permission from the Nature Research Ecology & Evolution Community.
Read the original paper here: What can horned beetles tell us about the mechanisms of plasticity and their evolution?
Interview with Sofia Casasa
Dr. Casasa, can you talk to us a bit about your research trajectory and how you came to settle on plasticity (and its evolution) as the main driving question behind your work?
I received my undergraduate degree from the UNAM (Mexico City), where I worked in a cell biology lab that focused on describing programmed cell death in rat ovary development. It was then that I started to become interested in development at the organismal rather than cellular level. In particular, I was fascinated by the role of the environment during development. For my PhD, the lab of Armin Moczek at Indiana University was the perfect place to start exploring multiple aspects of developmental plasticity. There, I was able to study the mechanisms of developmental plasticity and their evolution, as well as the role of plasticity in evolution.
What is particularly challenging about designing experimental systems to study plasticity?
One of the main challenges of studying developmental plasticity is that organisms that display the most diverse developmentally plastic phenotypes are typically emerging model systems. This poses multiple challenges that range from animal husbandry to molecular tools available. In our study, for example, we only had one of the species’ genomes available. This is more than many other emerging model systems have, but still posed challenges during our analyses.
This paper was based on the last chapter of your dissertation and we read it with great excitement!
I was very excited to have it come out too! It is great to hear that others are excited about my work.
What kinds of questions will your work open up for future study?
Our study was one of the first genome-wide characterizations of the transcriptional mechanisms of plasticity. One of our main findings was that the size of gene repertoire associated with plasticity mirrored the degree of plasticity at the morphological level. Future studies in additional systems with similar degrees of plasticity would add support as to whether this is a widespread feature of developmental plasticity. Our works also yielded a list of potential candidate genes to expand our understanding of the mechanisms of plasticity.
Additionally, we found that the evolution of novel degrees of plasticity was possible by a combination of taxon-restricted and conserved genes. An area of future research that I am particularly interested in is knowing how conserved, pre-existing gene regulatory networks are coopted into the evolution of novel plastic traits. For example, are some gene regulatory networks predisposed to be recruited into the evolution of plastic traits over other networks? And, how are taxon-restricted genes integrated in the network into a fully functional plastic response?
What will you be working on next? Are dung beetles still in the picture or can we expect more interesting models to analyze the mechanisms and evolution of plasticity?
I am currently a postdoc in the Ragsdale lab at Indiana University. I am using the lab’s model system, the polyphenic nematode Pristionchus pacificus, to better understand the mechanisms and evolution of plasticity. More specifically, my work seeks to understand the origin and evolutionary history of polyphenism genes across the nematode family (Diplogastridae), in which the polyphenism arose. By using genomics and transcriptomics I am investigating how genes in rapidly evolving gene families are integrated into ancestrally plastic networks. More on this to come out soon!
Affordances, or the possibilities for acting in our environments, are pervasive in everyday life. We are constantly surrounded by them: we perceive the floor as a walkable surface, a coffee mug is perceived as a graspable object, and doors are perceived as pass-through-able apertures, etc. For some authors, these objects of perception are, by far, the ones we interact with most throughout the day. If this is so, how is it that they are not deployed as commonly as other perceptual objects in the philosophical and scientific literature to make sense of our perceptual experience (compared to, say, colours)? Why are they scientifically understudied compared to other perceptual phenomena?
The answer is that, despite being so common, affordances have not been a widespread object of study in philosophy and behavioral sciences because they were not conceptualized as such until the second half of the 20th Century. Affordances were named by James Gibson, a pioneer in the establishment of ecological psychology. This discipline advanced, during the 1960s and 1970s, the main theses of situatedness, embodiment, and non-representationalism that are present in the post-cognitivist or embodied cognitive sciences born in the 1990s1. Decades before the establishment of the embodied and situated cognitive sciences, the ecological approach challenged key ideas of both behaviorism and cognitivism: for example, the understanding of perception as the passive reception of sensory stimuli, the thesis of the poverty of the stimulus, and the claim that psychological processes were based on mental representations, among others.
Contrary to these statements, ecological psychologists claimed that, in order to understand psychological processes, we should start not from the body or from the brain alone, but from the mutual and constant interaction of organism and environment (what they call the organism-environment system). This aspect is key and could be considered as one of the first attempts to establish a situated approach to cognition. They also claimed that organisms are active explorers of their environment (so perception is intimately connected to action), that organisms are constantly surrounded by information for action (a special kind of information that has nothing to do with information theory), and that the use, detection, or picking-up of that particular information is enough to explain how they perceive and act (no enrichment of the sensory stimuli is needed so as to form representations)2, 3, 4, 5, 6, 7, 8, 9, 10. In doing so, the ecological approach offers profound conceptual innovations, as it aims to overcome well-established dichotomies that have been present in the psychological and philosophical literature for centuries, such as mind-world, subjective-objective, and perception-action. Another distinctive feature of ecological psychology compared to current embodied and situated approaches is the vast corpus of in vivo experimental results (initiated since the 1950s and continued today) that support the theoretical principles of the discipline and that, among other things, prove the existence of affordances as objects of perception that can be scientifically described and studied11, 12, 13.
There have been several attempts to reunite affordances and evolutionary biology from different perspectives. Outside ecological psychology, authors such as Daniel Dennett (2017)14 have included affordances as key aspects to be taken into account when analyzing the evolution of cognition and consciousness. If we focus on those authors that related niche construction and affordances, it is worth mentioning that the project of Karola Stotz (2010, 2014)15, 16 aimed to develop an extended evolutionary psychology that includes niche construction theory and aspects of the embodied and situated cognitive sciences, incorporating the notion of affordance. Also, Denis Walsh’s (2014, 2015) project for naturalizing organismal agency from an extended evolutionary perspective includes affordances as an inspiration for understanding the environment. Within ecological psychology, several authors have always taken for granted that there is a strong link between both ecological psychology and evolution. Take, for example, the following quote from two of the main authors of this tradition:
Ecological theories not only assume that organisms exist in a rich sea of information about their environments, but also that they evolved in a rich sea of information. Consequently, it is supposed that the structure and function of the perceptual systems have become tailored to the available information. (Michaels and Carello 1981: 15)8
Despite the wide acceptance of the connection of ecological psychology and evolutionary biology, there has been no systematic research program to explain the intricacies of their entanglement in a detailed and meticulous way. Nevertheless, up to this date there have been two attempts at linking ecological psychology and evolutionary biology that are landmarks in the literature. The first and most important one has been Edward S. Reed’s (1996) approach to grounding ecological psychology in natural selection9. This has been the most ambitious and thorough approach to the issue to date. According to Reed, affordances exert selection pressure and organisms compete for those resources (Reed 1996: 26-27). Some authors within the ecological tradition criticized this approach, as according to their view it gave priority to the environment over the organism, and this is something that goes against the organism-environment mutuality emphasized by ecological psychology (Chemero 2009: 146, Withagen & van Wermeskerken 2010: 495). Following this line of criticism, Rob Withagen, Anthony Chemero, and others17, 18, aimed to reconcile ecological psychology and evolution by appealing to niche construction theory. When they contrast their views with Reed’s views, they claim that that both approaches (Reed’s and those of the authors that support niche construction) are incompatible, and that the connection between ecological psychology and evolutionary biology should be reconceptualized. In fact, they explicitly claimed that “the niche construction perspective suggests alternative roles [to the one offered by Reed’s project] for affordances in evolutionary dynamics” (Withagen and van Wermeskerken 2010: 497). As such, these two different ways of understanding affordances in the evolution of cognition (as elements of niche construction processes or as resources that exert selection on organisms) provide different understandings of the role of affordances themselves, and it has serious consequences for the role of agency and the role of the environment in evolution, leading to two irreconcilable and competing views about the evolution of cognition.
However, I think this is not the case. In a recently published paper entitled The evolutionary role of affordances19, I challenged this alleged incompatibility by showing that these two views are not competing but, rather, complementary. I proposed that these two different evolutionary mechanisms (natural selection and niche construction) can be understood as two different stages of the same continuous evolutionary process and that affordances play a different role in every stage.
Take, for instance, the well-known and intuitive example of a beaver population building a dam. As such, the environment, previous to be altered by the beavers’ action, affords certain actions and consequently exert some particular pressures (this actions can be either beneficial or harmful for the population’s chances of reproduction and survival). The construction of a dam is an alteration of the environment that also alters the available repertoire of affordances, not only for beavers themselves, but for the whole variety of living beings that populate the ecosystem, leading to unexpected consequences: with dams, beavers now have a place that affords hiding from predators and, given that dams alter the flux of the river, it also allows for the reproduction of certain microorganisms; it attracts migrating birds to stop nearby, and those birds also attract some predators, etc. As such, these alterations modify the affordances of the niches of all populations in that ecosystem and, with that, the selective pressures change with them. The offspring of those animals will receive a different environment compared to the one their ancestors found, one with in which these ecological inheritances will afford them different opportunities for acting that exert their own selective pressures. And so the story goes, cyclically. But the affordances that emerge will be, in different stages, either ecological inheritances of a niche construction process or selective pressures that count as key aspects in the process of natural selection of such population. As we can see, affordances have this double role in evolutionary processes, something that also shows how intricate are niche construction and natural selection. In fact, this example of the evolutionary role of affordances supports the idea that “[w]hen such modifications [of the niche] alter natural selection pressures, evolution by niche construction is a possible outcome” (Laland et al. 2016: 192)20.
If my proposal is on the right track, this can have consequences not only for starting to unfold the evolutionary role of affordances, but also for evolutionary biology as such, since they can help us understand in which specific sense these two evolutionary mechanisms are not isolated at all. This in turn might lead to a general, shared picture of the two previously supposed incompatible views. I develop this in detail in the publication.
Needless to say, this humble and little contribution to a detailed and complex debate that was initiated decades ago is far from fully unraveling the evolutionary role of affordances. Much more work is needed, both conceptual and empirical, for fully understanding the contribution of affordances in the evolution of cognition. I am sure that, as their contribution becomes clearer, it will also help the scientific and philosophical communities in their task of redefining the evolution of cognition under the principles of the Extended Evolutionary Synthesis.
Read the original publication here: Heras-Escribano, M. (2020). The evolutionary role of affordances: ecological psychology, niche construction, and natural selection. Biology & Philosophy, 35, 30.
1.Heras-Escribano M. 2019. The philosophy of affordances. Cham: Palgrave Macmillan.2. Gibson JJ & Gibson EJ. 1955. Psychological review.3. Gibson JJ. 1966. The Senses Considered as Perceptual Systems. Boston: Houghton Mifflin.4. Gibson EJ. 1969. Principles of perceptual learning and development. New York: Appleton-Century-Crofts.5. Gibson EJ. 1991. An odyssey in learning and perception. Cambridge, MA: MIT Press. 6. Gibson JJ. 1979. The Ecological Approach to Visual Perception. Boston: Houghton Mifflin.7. Turvey et al. 1981. Cognition. 8. Michaels CF & Carello C. 1981. Direct perception. New York: Prentice-Hall.8. Reed ES. 1988. James J. Gibson and the psychology of perception. Yale University Press.9. Reed ES. 1996. Encountering the world: Toward an ecological psychology. Oxford University Press. 10. Heft H. 2001. Ecological psychology in context: James Gibson, Roger Barker, and the legacy of William James’s radical empiricism. Psychology Press.11. Richardson et al. 2008. in Handbook of cognitive science: an embodied approach.12. Chemero A. 2009. Radical embodied cognitive science. Cambridge: The MIT Press.13. Turvey MT. 2018. Lectures on perception: An ecological perspective. Routledge.14. Dennett DC. 2017. From bacteria to Bach and back: The evolution of minds. WW Norton & Company.15. Stotz K. 2010. Phenomenology and the Cognitive Sciences.16. Stotz K. 2014. Frontiers in psychology. 17. Withagen R., Chemero A. 2009. Theory & Psychology.18. Withagen R. & Wermeskerken M. 2010. Theory & Psychology.19. Heras-Escribano. 2020. Biol & Phil. 20. Laland KN et al. 2016. Evolutionary Ecology.
Tobias Uller writes:
West-Eberhard famously suggested that plasticity ‘takes the lead’ in adaptive evolution. But how can you tell if you are not there to see it happen? In a new paper in Evolution Letters, we show one way to tackle the problem. We figured that we can look for signatures of plasticity by comparing the magnitude and direction of plastic responses to the environment with how locally adapted populations differ.
The literature on plant local adaptation is full of reciprocal transplant experiments that record both plasticity and adaptive divergence. We therefore used such data to test a number of hypotheses – and here is what we found:
First, the ancestral plastic response was generally well aligned with the difference between locally adapted phenotypes, making plasticity appear to ‘take the lead’ in adaptive evolution. Or – if you prefer – the plastic response of ancestors left a signature in the locally adapted phenotypes of descendants.
Second, plastic responses were sometimes more extreme than the locally adapted phenotypes. This can give the false impression that plasticity and evolutionary adaptation oppose each other: in fact, truly maladaptive plasticity was rare.
Third, although the signature of plasticity generally persists during local adaptation, genetic evolution modified trait combinations independently of plasticity.
We learnt a lot during this study. Perhaps the most important message is that we are a long way from understanding the relationship between plasticity and evolution – largely because very few studies are actually designed to test if evolution goes where plasticity leads.
Hopefully this paper shows not just that this is a hypothesis that can be tested, but also help to clarify what kind of data that we need.
Publication:
We have just launched a new resource website on niche construction and niche construction theory: nicheconstruction.com
This website was developed in the hope that it will prove a valuable resource for the community of academics, students and other individuals interested in the topic of niche construction. The website is designed to provide an accessible introduction to the subject of niche construction, explaining what it is, and how it is being used across multiple academic fields.
There are so many published articles on niche construction now that we cannot possibly feature them all. Please forgive us if we have not mentioned your favourite study. Designing and producing the content for this website has been a huge undertaking, and the website is already far more content rich that most comparable sites. Nonetheless, if you do detect an egregious omission please don’t hesitate to let us know, and maybe we will be able to incorporate it.
If you have recently written on niche construction, tell us about your work, and we will try to tweet about it from @EES_update. Our hope is that a flavour of what is genuinely exciting about niche construction comes across in the website’s pages, that it starts to become widely used, and eventually becomes cherished by the community.
Kevin Laland and Lynn Chiu
May 2020
Darwin was particularly fascinated by changes in animals that occurred during domestication which helped to form some of his major ideas. Indeed, the Origin of Species contained a chapter on pigeons, and Darwin even bred pigeons himself to understand the processes taking place during domestication. However, despite this long-standing interest in domestication evolutionary biologists are still unsure about how it is initiated.
Outside of domestication we now understand that humans apply significant selective pressures on wild populations and even evolutionary change. While often unintentional, such selection has led to changes in wild populations such that animals adapt to the presence of humans and the resources they provide. For example, animals are moving into cities. Urban ecology, in particular, has emerged as an important sub-field for understanding how wild populations interact with and persist in urban settings. Increasingly, findings indicate that urban habitats are imposing selection pressures that differ substantially from rural habitats (Schilthuizen 2018). These differences are reflected in various phenotypic traits that change between urban and rural habitats.
We were interested in three basic questions: 1) could these phenotypic changes related to urban habitats be useful for informing us about how domestication processes are initiated? 2) could current selection pressures differ between urban and rural habitats to explain these differences, and 3) could mechanisms for how variation arises (in complement to selection) be part of how divergence between urban and rural habitats occurs?
Although in many cases a heritable basis for changes between urban and rural habitats needs to be verified, they are often quite fast. We frequently think that this is just the nature of contemporary evolution, whereby selection can rapidly do as it pleases. However, perhaps these responses are aided by past evolutionary experiences for a given lineage. It would seem very unlikely that a lineage would, or even could, completely dispose of the machinery that has evolved for adaptation in past environments (Parsons et al. 2020).
It would therefore seem that developmental biases in how variation arises could also be influencing these outcomes to human-induced selection by limiting the range of possible phenotypes available. In other words, the phenotypic variation available could be primed for rapid evolution by such biases that actually speed the rate of change (Uller et al. 2018). Past selection would have favoured phenotypes adaptive for past environments and the developmental machinery to put this in place may sometimes be re-utilized in many cases of rapid evolution. But could these rapid responses to human environments also be related to the initiation of domestication? It takes a special animal to address both of these questions together.
Our current study aimed to address these issues using comparisons of urban and rural populations of red foxes (Vulpes vulpes). Red foxes have become extremely common in urban habitats over the past century, especially so within the UK where they are often viewed as a pest species. Urban lifestyles are quite different for foxes causing them to shift diets toward our wasted foods, while also causing a substantial reduction in the size of their territories. From an urban ecology perspective this provided an opportunity for study as skulls from foxes that were culled from central areas of London, as well as the surrounding countryside, were readily available.

Photo by Sean Page
However, foxes were also already an especially important animal for our understanding of domestication processes. In Russia, the famous Belyayev domestication experiment on foxes has been running since 1959. This project has solely favoured behavioural ‘tameness’, or more precisely ‘friendliness’, towards humans and was intended to mimic the hypothesized selection regime during the initial domestication process of dogs. Despite this focus on behaviour, it has resulted in a number of morphological changes including a relative shortening of the snout, novel coat colours, and reduced brain size. These trait changes follow predictions from ‘domestication syndrome’ whereby suites of traits are commonly found to change under the process of domestication.
Our focus for red foxes was on their skull morphology and this allowed us to address some of these traits, namely the shape of the snout and the size of the brain case (indicative of brain size). We found that snout length was reduced, along with the brain case in urban foxes relative to their rural counterparts (Parsons et. al. 2020). Also, we found that the degree of sexual dimorphism was reduced in urban foxes. All three of these findings strikingly matched with patterns found within the Belyayev experiments and was in alignment with domestication syndrome.
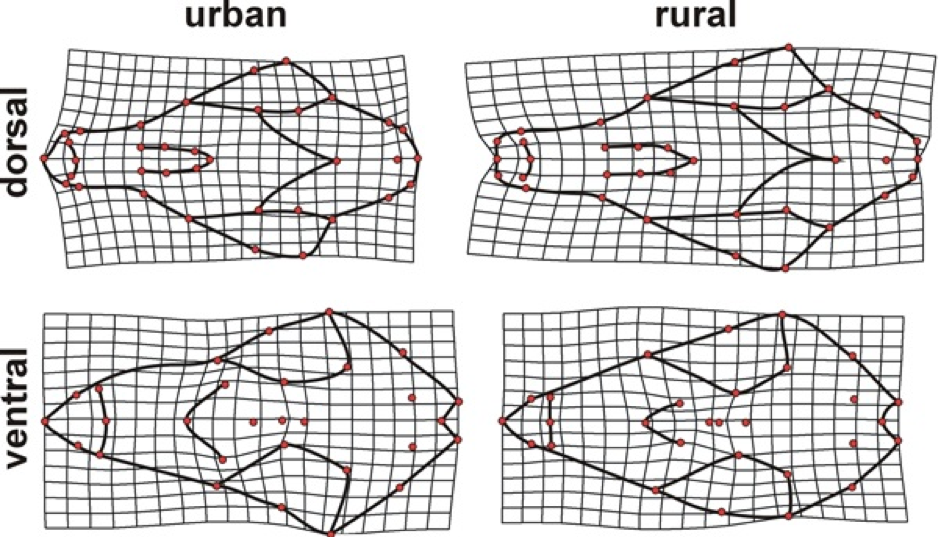
Shape differences between urban and rural foxes were especially notable in the width and length of the snout. Urban foxes had a shorter somewhat wider snout relative to rural foxes, while there was also a slight reduction in the size of the brain case. Deformation grids depict shape variation between urban and rural fox skulls magnified by 3X in the dorsal and ventral aspects.
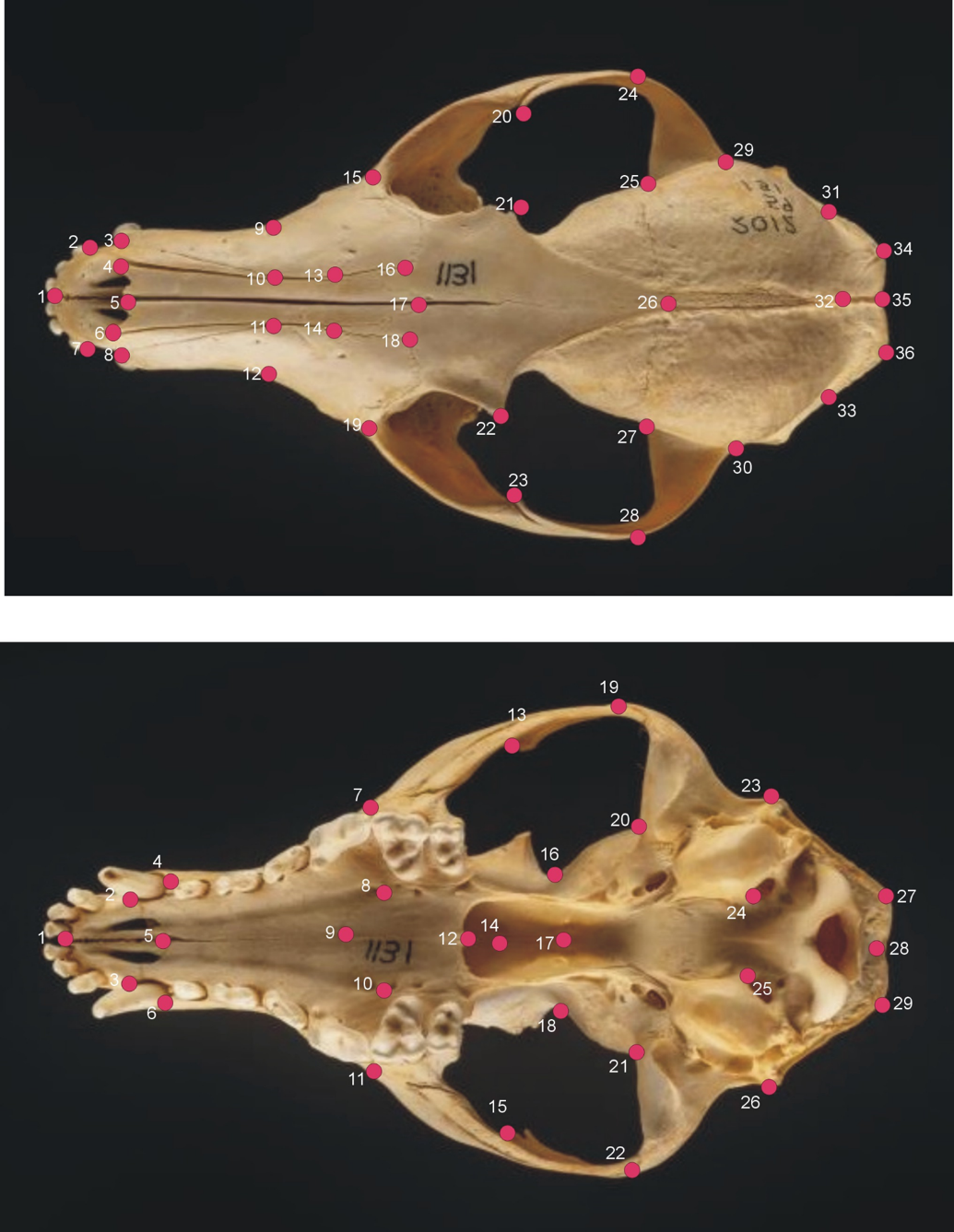
The dorsal and ventral aspects of red fox skulls depicting the landmarks in red we used to measure geometric shape.
While we couldn’t directly test for adaptation, these differences provided biomechanical interpretations. For example, a shorter snout would confer a slower bite, but one with a higher bite force. This may be an advantage in urban environments for gaining access to various stationary foods. A faster jaw would likely help more in rural habitats where there would be a stronger reliance on motile prey. Brain size differences need further investigation, but the reduced size of the brain case in rural foxes may reflect a reduced cognitive demand from living within a smaller territory.
As mentioned, we wanted to know whether these patterns of evolution reflected deep-seated developmental biases. So we looked at skull morphology across other canids, mostly from within the Vulpes genus. These species have been separated for millions of years and reflect a range of ecology. To our excitement it turned out that while the magnitude of differences was several folds greater across these species, the way in which they were divergent was the same! In other words, the main type of variation that differed across species involved a shortening of the snout accompanied by a reduced brain case, or a lengthening and narrowing of the snout accompanied by an increase in the brain case. This similarity in the patterns of divergence was suggesting to us that developmental biases have been operating in foxes for millions of years, and continue to operate in what is very likely a case of rapid evolutionary divergence between urban and rural habitats. In this sense developmental biases could provide a direct link between micro- and macroevolutionary divergence. Simultaneously, these findings also suggest that developmental biases can help to initiate the changes that prime some animal for domestication. But what mechanisms could be linking these patterns of evolution?
For domestication syndrome, Wilkins et al. (2014) proposed that a link among the many traits involved may lie in their cellular origin as they all appear to be derived from neural crest cells (NCCs). NCCs are a vertebrate specific cell population, sometimes argued to be a fourth germ layer, that arise during early embryogenesis at the dorsal edge (crest) of the neural tube. These cells migrate throughout the body toward the cranium and trunk and provide the cellular precursors of many cell and tissue types, including many of the bony elements of the skull, elements of the brain and nervous system, the adrenal gland, and melanocytes. Wilkins et al. (2014) present extensive data showing strong similarities between the traits associated with domestication syndrome and the phenotypes produced by loss-of-function mutations in neural crest cell genes. The same genes are likely candidates for the genetic changes underlying both the shift from rural to urban living, and the similarly biased pattern of diversification amongst canids. Many of the genes involved in the genesis and migration of NCCs have been well characterized and should be considered targets for future genomic exploration and functional genetics.
More broadly, this study demonstrates that when we examine multiple levels of phenotypic divergence, even the most rapid types of contemporary evolution can be connected to long-term evolutionary trends. Developmental bias may offer a perspective that helps make understanding these patterns tractable across a range of organisms. First, it can focus interest on what phenotypes arise, and or, can’t arise over evolutionary time. Second, given what we know about the molecular and cellular basis of morphological development it can also motivate studies that can be translatable across taxa. For example, finding a molecular or developmental ‘waypoint’ for bias in foxes may lead to a deeper understanding of evolutionary patterns across canids and many other vertebrate families.
Key references:
Schilthuizen M. 2018. Darwin comes to town: how the urban jungle drives evolution. Quercus: London
The logic underlying cell differentiation has motivated an intense field of debate over years. How can plastic, developing cells “know” exactly where and how to differentiate ? Given that cells are aquipped with genetic networks, could they benefit from some form of basic learning, as cognitive systems do with neural networks? In our recent paper, we show that this idea goes beyond a simple metaphor: differentiating cells can exhibit basic learning capabilities that enable them to acquire the right cell fate. Although this statement could sound odd to some, we show how such a learning ability naturally arises from the way in which plastic cells integrate different signals from their environment during development. But to better grasp this connection, we have first to say a few words about the phenotypic plasticity of cells.
Phenotypic plasticity (i.e. the sensitivity of an organisms to its environment) is pervasive at every level of the biological hierarchy. For example, whilst part of the leaf morphology in plants is specific of each species, a great amount of variation in leaf size and shape can be linked to specific variations in the light, moisture and temperature conditions of the local environment. For those familiar with the topic, it is common to conceptualize plasticity as a “reaction norm”: an abstract function that link a specific environmental input to a phenotypic state. These reaction norms are often assumed to be genetically encoded and simple (e.g. linear) functions of a single environmental cue, so that phenotypes vary gradually in response to changes in one environmental variable (Fig. 1A).
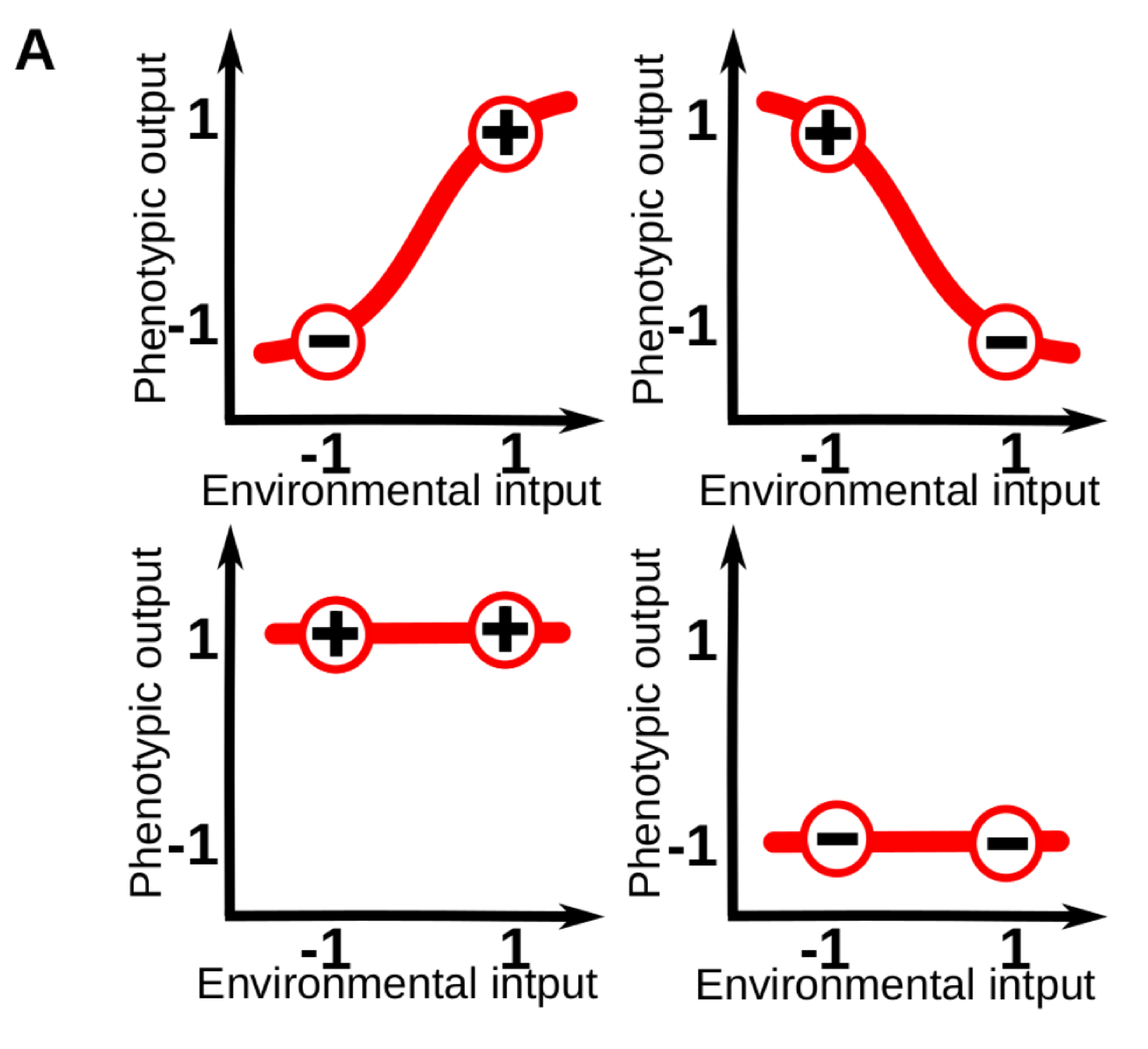
Figure 1A. Models of phenotypic and cell plasticity often depict a 1-dimensional reaction norm (red line), for a single continuous environmental cue.
Classic reaction norms are often a good approximation, but in reality the phenotypes are commonly determined by many environmental cues acting simultaneously. This is very relevant in the case of cell differentiation in multicellular organisms, where developing cells acquire their final fate by integrating several informational inputs from their surrounding tissue micro-environment (Fig. 1B).
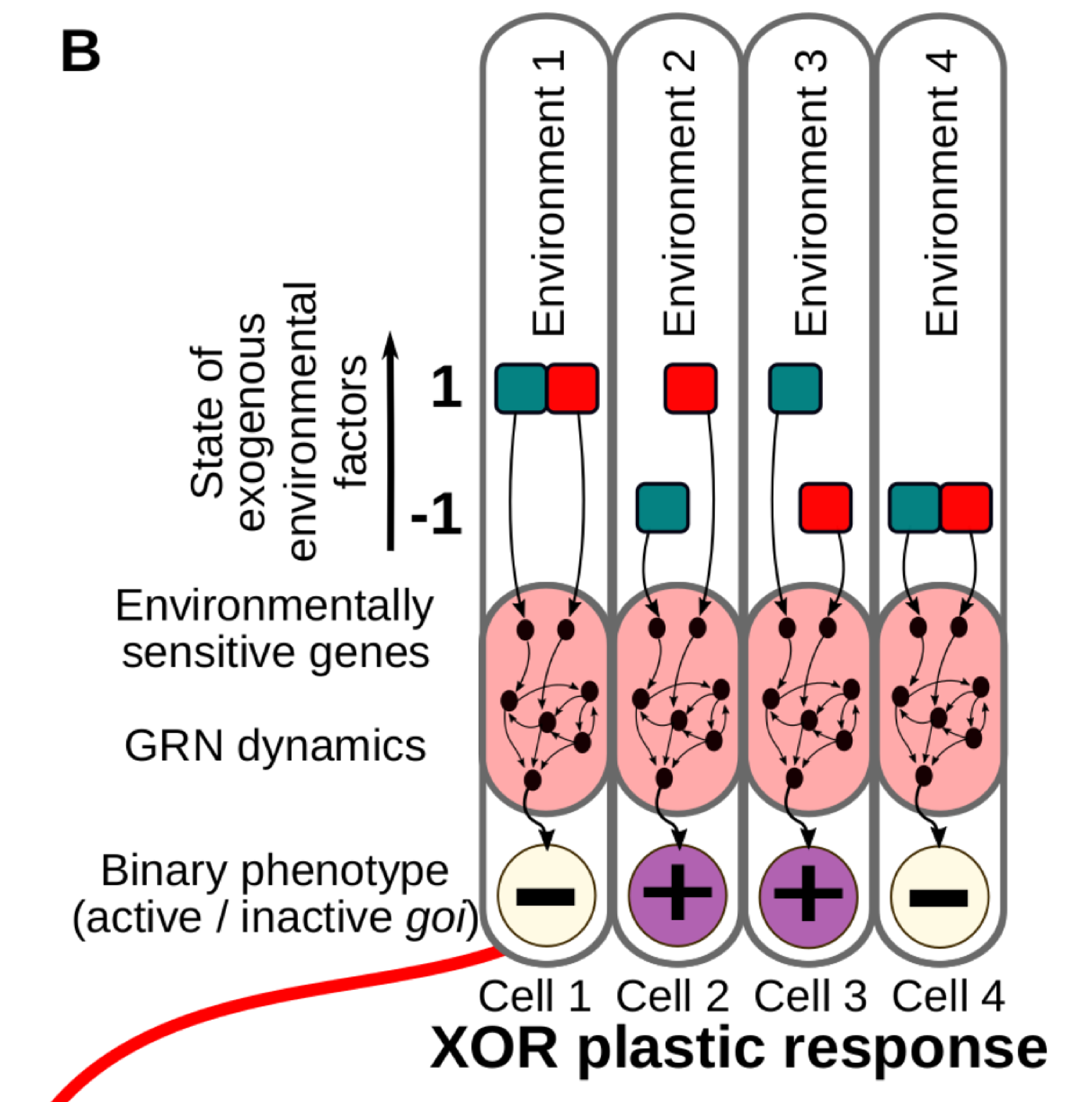
Figure 1B. We consider different combinations of Ne discrete environmental factors (in this illustrative example, Ne = 2, red and blue in the figure) determining a binary cell state. In this case, a number of possible environment-phenotype interactions can be described by means of logical functions derived from Boolean algebra (right).
These inputs include signalling molecules secreted by other cells, nutrients and chemicals taken up from the external environment, and even interactions between the embryo and other organism, such as the bacteria colonizing its skin and gut. Furthermore, the relationship between the environmental cues and the phenotypic responses can be highly non-linear. This calls for representations of plasticity in terms of “multi-dimensional” reaction norms, but these remain largely unexplored.
This work provides new conceptual tools to characterise these “multi-dimensional” reaction norms and to better understand how they arise and evolve, especially in the context of cell differentiation. In a nutshell, the relationship between the environmental cues (inputs) and cell phenotype (output) is described using Boolean logical functions (Fig. 1C).
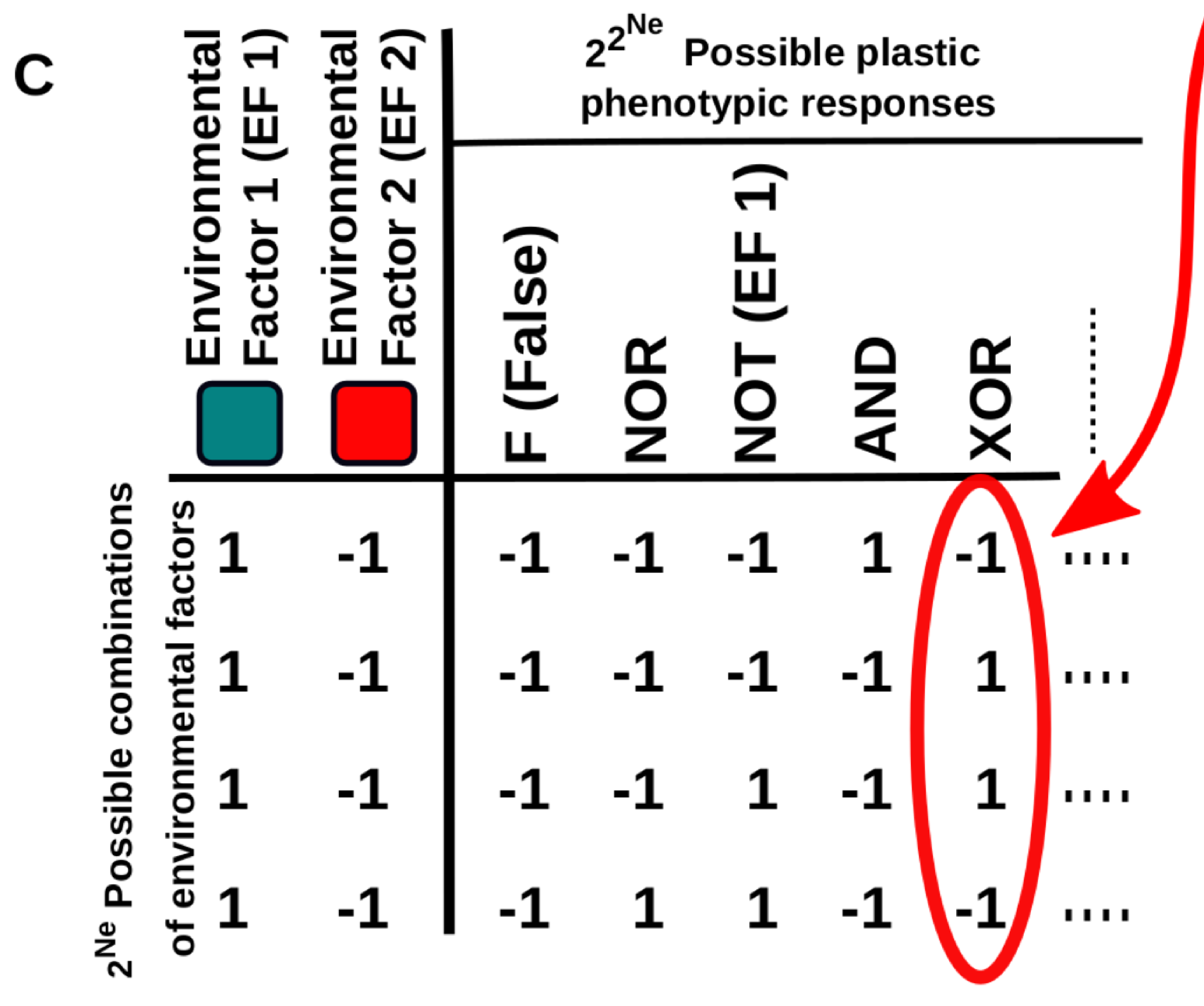
Figure 1C. Five examples of multi-dimensional reaction norms in which the expression level of some gene is affected by the Ne environmental factors. One of these multi-dimensional reaction norms (e.g. the one described by the “XOR” logical function depicted in B) is set as a target of a continuous GRN-based model.
This representation is very useful because it naturally introduces a way to measure the complexity of a multidimensional reaction norm: it will be very simple if the cell state is determined by one of the inputs, a little bit more complex if it is determined by a linear combination of the inputs (e.g. cell gets the state “A” if both inputs are negative and “B” otherwise) and maximally complex if the cell state is determined by a non-linearly decomposable function of the inputs (e.g. cell gets the state “A” if both inputs are equal -positive or negative- and “B” if they are different). In addition, this abstraction allows us to apply principles derived from computer science and learning theory to the study of cell plasticity.
By using a biologically realistic model of gene regulatory networks (GRNs), we show that natural selection is capable of discovering many forms of cell plasticity, even those associated with complex logical functions (Fig. 2B).
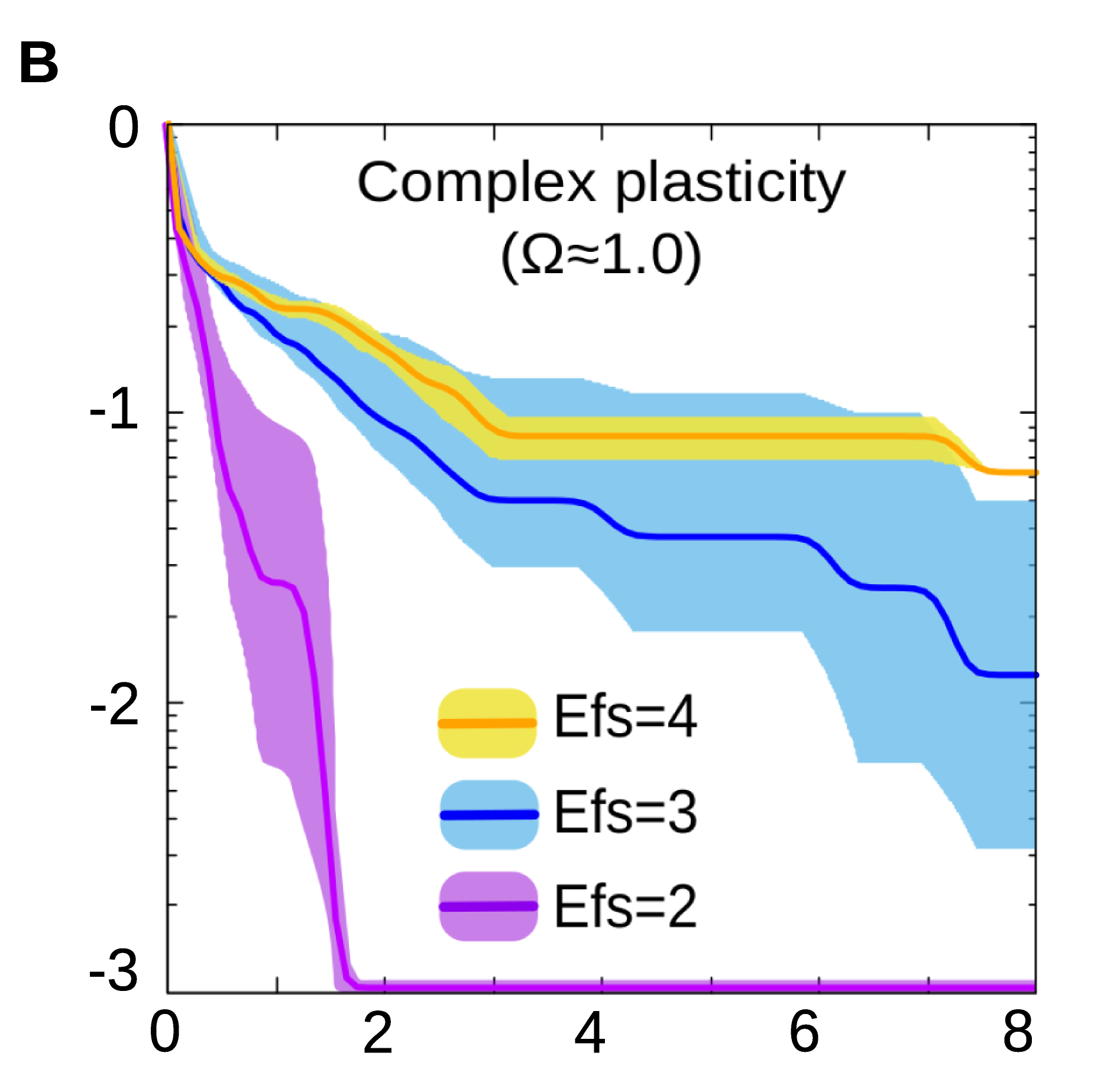
Figure 2B. In general, evolving a specific form of cell plasticity is faster when the number of environmental factors involved is low.
These later would correspond to scenarios where the cell state depends on the simultaneous integration of several environmental signals, but none of these signals alone contains enough information to determine the response (e.g. a response which is triggered if all the inputs are equal will require to know the state of every input, from the first to the last one).
This capability of plastic cells to evolve complex reaction norms is even more patent when the environmental signals modify the strength of regulatory interactions between genes. To illustrate this, take the phenotypic effect of temperature. Temperature does not to directly enhance or repress the expression of a gene but affects the kinetics of interactions between different gene products (Fig. 3A-B).
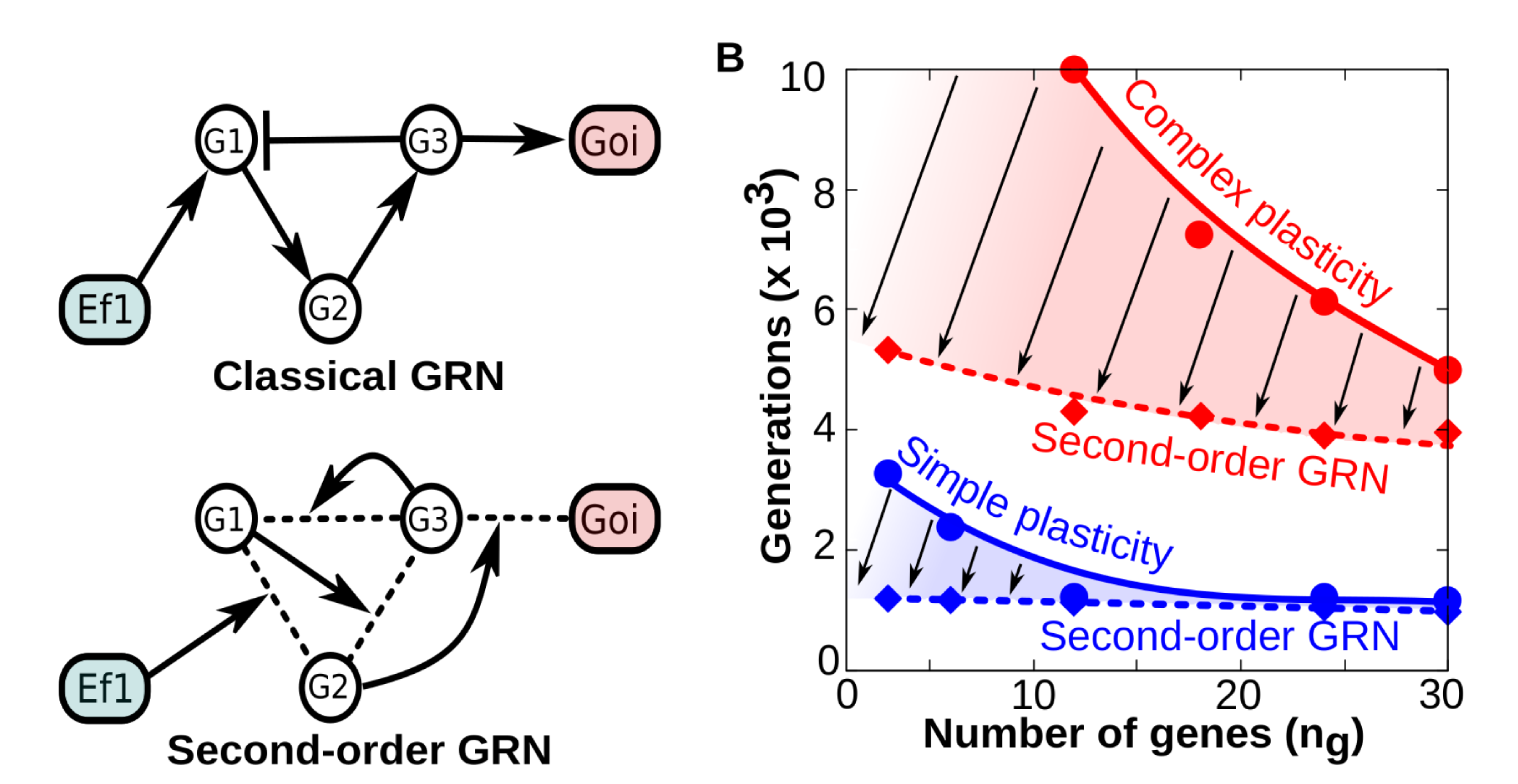
Figure 3A-B. A) Contrary to “classical” GRNs in which the network topology remains fixed over developmental time, second -order (tensor-based) GRNs have dynamical topology: each gene-gene interaction strength is determined by the concentration of other genes (g1, g2, etc) and environmental factors. In both cases the final phenotype is recorded as the binary state of a gene of interest (goi). B) Comparison between “classical” and “second-order” GRNs in terms of their evolvability (how many generations they need to achieve a fitness W≥0.95 when they are selected to represent specific reaction norms). The panel shows that everything else being equal (number of genes, function complexity), GRNs with dynamical topologies (dashed smoothed lines) require much less generations to evolve any reaction norm than classical GRNs (solid smoothed lines). Thus, GRNs capable of second-order dynamics exhibit higher evolvability than classic GRNs (being the difference more patent for complex forms of plasticity).
Simulations also reveal that developmental dynamics produces a strong and previously unnoticed bias towards the acquisition of simple forms of cell plasticity. This bias causes linear reaction norms to be far more likely to appear than humped-shaped or sigmoidal ones, and appears even in random (i.e. not evolved) GRNs. This suggests that this bias towards simple plastic responses is an inherent feature GRN dynamics rather than a derived property (Fig. 4A).
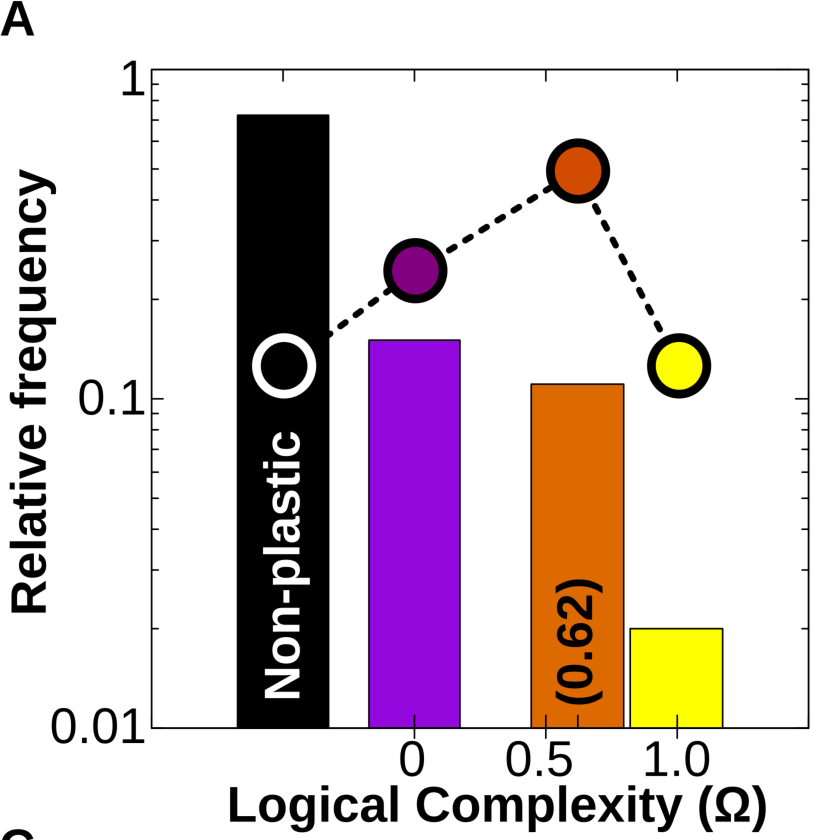
Figure 4A. (Log) Relative frequency of different types of cell plasticity according to their complexity Ω in a vast GRN space. Black column: no plasticity (the phenotypic state is purely determined by genes); purple column: phenotypic state directly determined by just one of the Ne environmental factors; orange column: phenotypic state determined by simple combinations of the Ne environmental factors (linearly decomposable functions) and yellow column: complex forms of cell plasticity associated with non-linearly decomposable functions (XOR, XNOR; see SI). Dashed line and dots represent the relative distribution of each family of logical functions in the mathematical space. We see that although the number of simple and complex functions that exist is approximately equal, GRNs produce simple functions much more often.
In a second set of experiments, we explored the evolutionary consequences of that bias. These experiments show that, when the selective environment mirrors the plastic bias of development (that is, when simple reaction norms have higher adaptive value), plastic cells are able to display appropriate plastic responses even in environmental conditions that they have never experienced. This ability to generate variation is a form of basic learning. New variation is not produced at random, but in an advantageous direction (Fig. 6A-B).
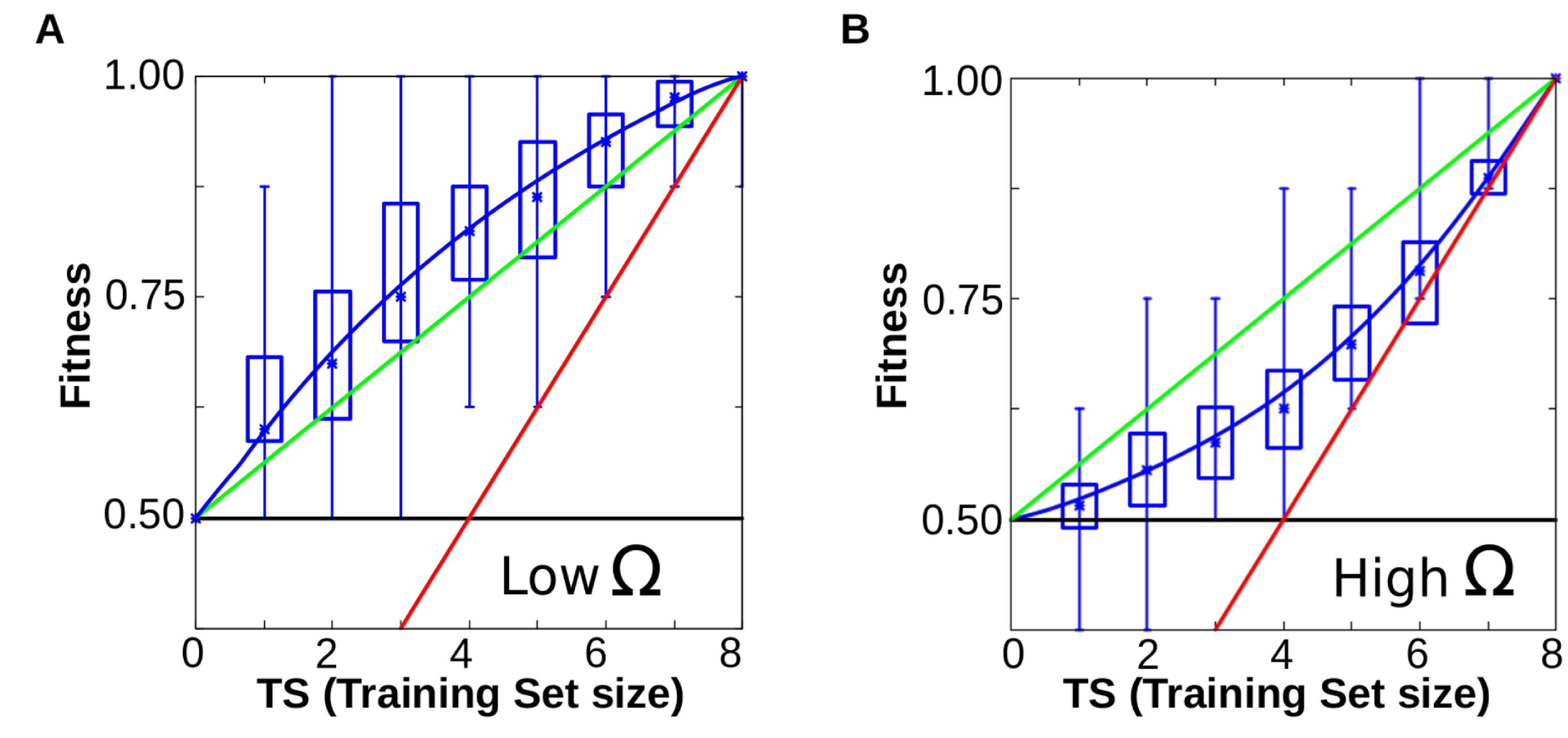
Figure 6A-B. When cells evolve simple forms of plasticity, they are able to generalise, performing better than chance in previously unseen environments (red line represents the information provided (x = y), green line represents the expected performance at random (2Ne-TS)/2; and blue line the degree of matching between the resulting phenotypes and the expected ones for a given function). Notice that blue line runs consistently higher than the green line (random response). B. Similar experiments yield poor performances when cells have to evolve complex forms of plasticity (blue line below the green one, see main text).
As a proof of principle, this work shows how differentiating cells can take advantage of such basic learning to acquire the right cell fate even in noisy developmental conditions (Fig. 7).
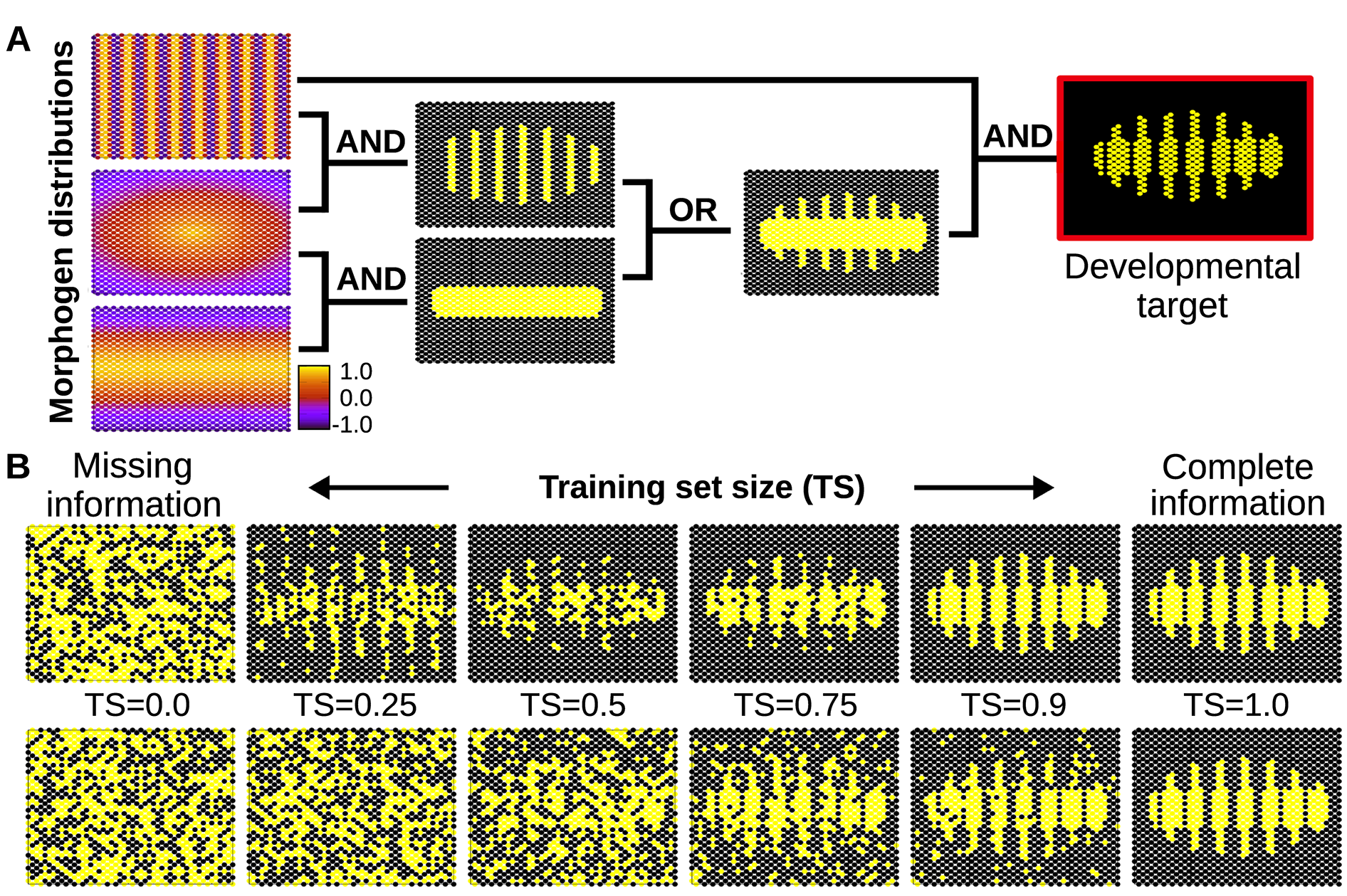
Figure 7. A) Simple spatial distribution of three morphogens over a two-dimensional (50×25) field of cells (similar gradients are commonly found in the early developmental stages of many organisms). In this example, these three environmental signals are integrated by individual cells (according to relatively simple logical functions) to form a segmented developmental pattern. The complete plasticity function is represented by a truth table of 32 rows specifying whether the relevant cellular output should be on or off for each possible combination of morphogens (see Methods). B) Individual cells are evolved under selective conditions that expose them to some rows of this table but not all. Under this scenario of missing selective information, cells equipped with real GRNs perform by default a simple logical integration of the morphogen cues, which in this example results in a much more robust and uncertainty-proof developmental process (upper row). When the training set contains all the information necessary to completely define the plasticity function, natural selection finds a GRN that calculates this function accurately (right). Of course, when past selection contains no useful information, the phenotype of the GRN is random (left). In between we can see that the generalisation capability of the evolved GRN ‘gives back more than selection puts in’- i.e. the phenotype produced (top row middle) is visibly more accurate than the training data experienced in past selection (bottom row middle). This is quantified in S1 Fig. In the bottom row, cells do not exhibit any bias towards simple functions, and therefore they acquire any random form of plasticity compatible with the previously experienced environmental inputs. This randomness in the plastic response fails to fit the required (target) function, preventing generalisation (i.e. the system needs to evolve in a full-information scenario to produce the required phenotype, see main text).
If the developmental pattern emerges from a simple integration of many signals, the cells do not need to receive every signal to trigger the appropriate response: they will unleash a complete and adequate response from just a few inputs. The cells will most likely provide the right answer because they do not consider the whole, vast space of possible responses, but just a few simple, adaptive solutions compatible with the few bits of environmental information received. Metaphorically, this could be seen as if plastic cells would able to apply a sort of Occam’s razor parsimony principle to the developmental problems they must solve. This finding identifies a novel mechanism that increases the robustness of developmental process against external perturbations.
Overall, this work illustrates how learning theory can illuminate the evolutionary causes and consequences of cell plasticity. This approach is possible because learning principles and logical rules are not substrate specific: they can be exhibited by interacting molecules, genes, cells, neurons and transistors alike as long as these elements have the right type of interactions. This substrate independence of learning principles helps interpret the main finding of this work: that evolving gene networks can exhibit adaptive principles similar to those already familiar in cognitive systems.
Read the original article here: Brun-Usan M, Thies C, Watson RA. 2020. How to fit in: The learning principles of cell differentiation. PLOS Computational Biology. 16(4):e1006811.
Philosophers of science are sometimes seen by practicing scientists as useless, or unnecessary. Take scientist, writer, and broadcaster Neil deGrasse Tyson. Tyson recently poked fun at philosophical questions and methods, wondering whether a philosopher could ever be a “productive contributor to our understanding of the natural world.1” And many philosophers of science are familiar with the famous quote, usually attributed to Richard Feynman, that “philosophy of science is about as useful to scientists as ornithology is to birds.2”
Yet if this is the prevailing attitude among scientists, it certainly isn’t true for those working on the Extended Evolutionary Synthesis (the ‘EES’). Not only are key members of the project themselves philosophers, but leading scientists exploring the EES often co-author papers with philosophers, publish in philosophy journals, and employ philosophers on their collaborative projects. And I’m grateful for it. Until quite recently, I was employed through one of the large grants associated with the EES.
Combining philosophical and the biological sciences methods and tools can be both conceptually and empirically fruitful. Looking beyond the EES, one can point to the excellent interdisciplinary work coming out of Ford Doolittle’s or Thomas Pradeau’s groups, the productive collaboration between Kim Sterelny and Kevin Laland, the powerful game-theoretical models of Brian Skyrms, or the wealth of recent papers exploring finite population models. Philosophers of science—particularly philosophers of biology—have a lot to offer the biological sciences, and vice versa.
Still, when disciplines come together to work on interdisciplinary projects—whether this means philosophers writing on aspects of scientific theory or practice, or when scientists write philosophical papers—the backgrounds of these individuals’ training can become relevant. The kinds of questions philosophers have historically asked influence what they research now. In just the same way, the modelling frameworks, benchtop tools, and model organisms of scientists influence what phenomena they are exploring now and how they do so.
In a recent paper of mine (‘Synthesising Arguments and the Extended Evolutionary Synthesis’), I argue that an important feature of the debate around the EES results from some insalubrious mixing of philosophy and science. One of the reasons that hearts get pumping at the mention of the EES is that both proponents and skeptics alike have tended to frame the reception of EES ideas in terms of theoretical change, and sometimes, revolution.
For a sizeable chunk of the Twentieth Century, a dominant issue in the philosophy of science was the nature of scientific theory. After all, what are scientists up to all day, if not fretting about and elaborating their theories?3 Some of the more familiar names from mid-century philosophy of science worked on this question, like Karl Popper and Thomas Kuhn. A key project within this literature concerns how theory changes—how scientists decide to abandon one theory and move onto another. We’re probably all familiar with Popper’s heroic (but ultimately, unsuccessful) defense of falsification4, or Kuhn’s articulation of paradigms, revolution, and pre-paradigmatic science, but these are just two positions among many put forward at this time.
Are the language and tools of theory-change useful for understanding whatever proponents of EES are up to? As I said, writers both optimistic and skeptical about the merits of the EES often deploy the language of theory-change, and acknowledge writers like Kuhn or Imre Lakatos. This is so even when they deny that evolutionary theory needs anything as radical as a Kuhnian paradigm shift.
But a hazard of deploying this language is that it appears to treat the EES as comprehensive, full-scale alternative to the theoretical resources at work in contemporary biology5. And this, I argue, is not the case. The EES just isn’t denying the foundations of evolutionary theory in a way that might license talk of theory change. Worse, using such language obscures how we really should understand the EES and evaluate it.
The positive story I put forward builds on the work of philosophers Alan C Love, Ingo Brigandt, and James Griesemer6. The basic idea is that evolutionary theory itself can be mobilized and represented in various ways. Think of the way that the same land area—say, Banff National Park—can be represented on a map. One map might highlight roads and access ways. Another, the location of recent Grizzly activity. A third, major geological strata. And so on. Each map covers the same terrain, but selectively highlights features of it in ways that can facilitate different projects (driving, avoiding bears, finding hot springs, etc.). In the same way, I argue that the EES itself mobilizes the tools, concepts, and methods of evolutionary theory in one way, while contemporary consensus methods mobilize it in another.
This is important, because on this understanding, theory change isn’t at issue. Both prevailing practice and the EES are using the same evolutionary theory, but adopting different parameterizations, boosting the importance of different processes, and occasionally mobilizing different tools, methods, and concepts. They’re making different maps of the epistemic terrain. What is important on this revised picture is how good those maps are—that is, how they help to answer specific research questions. Is non-genetic inheritance important to explaining evolution? Should we take reciprocal causation into consideration in our models? Well, it all depends on what we’re trying to find out about the world.
Looking at specific research aims—organized into what Brigandt and Love call ‘problem agendas’—helps us to see that in many places, the EES and current evolutionary methods may not be in conflict. Either the EES adopts the same kinds of tools as current practice, or is trying to answer a question that isn’t on their radar. But in some places, both the EES and current practice do try to address the same research question, and mobilize very different tools and concepts to do so. It is here, with these situations, where the EES and prevailing methods in contemporary biology can come into productive, empirically fruitful conflict.
It takes a lot of work to identify these areas, and make the differences between the EES and prevailing practice clear. In the paper, I focus on niche construction, and suggest that issues around evolvability might be where niche construction tools and concepts conflict with those of ‘standard’ evolutionary theory. But I think the effort is worth it. In the end, this approach better makes sense of what the EES is up to, how we should understand what skeptics and optimists are debating over, and how proponents should defend their efforts going forward.
Footnotes:
1. See the discussion and partial transcript at https://io9.gizmodo.com/1575784094
2. I cannot find an actual reference of such a quote, and in other places the quote is attributed to an anonymous source.
3. Quite a lot, as it turns out.
4. Why was Popper’s account unsuccessful? I suggest Tim Lewens’s (2015) The Meaning of Science for an approachable and balanced take on Popper’s philosophy.
5. This isn’t to say that such language is always problematic. Philosophers continue to explore issues raised by Kuhn and Lakatos, often to examine very general features of confirmation, theory-choice, and the development of scientific theories.
6. Brigandt and Love, ‘Conceptualizing Evolutionary Novelty: Moving Beyond Definitional Debates.’ J Exp Zool (Mol Dev Evol) 318 (2012):417-427; Griesemer, ‘Presentations and the Status of Theories.’ PSA: Proc Bienn Meet Philos Sci Assoc (1982):102-114.
Massimo Pigliucci, Tim Lewens, Zanna Clay
The brutal mechanism of natural selection gave rise to all life on our planet, or so the Darwinian story goes. But from mice to great whales, it now appears that animals can inherit socially acquired characteristics. And after a century of hype, the tools to build a brave new world through intelligent design are within our grasp. Is the theory of natural selection a Victorian relic, about to be replaced by a new science of cultural evolution? Or does the promised paradigm shift provide no real challenge to Darwin’s theory?
The Panel: Philosophers of science Massimo Pigliucci, author of Cultural Evolution Tim Lewens, and psychologist and primatologist Zanna Clay ask if there is more to evolution than we might think. David Malone hosts.
External Link: https://iai.tv/video/after-darwin
We are delighted to announce that the full volume of the special issue “Developmental Bias in Evolution,” edited by Armin P Moczek, is now out in the journal Evolution & Development. This special issue follows our third workshop at Santa Fe in 2018.
In November 2018 the Santa Fe Institute hosted a two‐day workshop titled Developmental Bias and Evolution, funded by a grant from the John Templeton Foundation. Involving 34 participants and 22 talks, the workshop covered a wide range of approaches toward the study of bias exerted by developmental systems in the production of phenotypic variation, the impact such bias might have on evolutionary dynamics, and the methods that exist to assess the nature and consequences of this impact. Talks included historical retrospectives, philosophical examinations, and a great diversity of empirical treatments of the subject. Significant discussion and debate followed each presentation, and creative tensions emerged around key issues that characterize the diversity of perceptions of what, exactly, constitutes bias in developmental systems, when or how such bias may be evolutionarily relevant, and at the most basic level, whether the concept of developmental bias is itself useful in fueling a productive research program. This special issue is meant to capture this diversity of viewpoints, and to provide a collection of perspectives that will inform and motivate the next round of research, and the next generation of researchers. — Moczek’s editorial, p.3
You can access the full issue here: https://onlinelibrary.wiley.com/toc/1525142x/2020/22/1-2